Research Topics: Behavioural and Systems Neuroscience, Epilepsy and Seizure Disorders, Neurobiology of Motor Maps, Learning and Memory.
My research program focuses on two independent but related fields of research that have both basic (curiosity driven) and applied (health sciences) aspects to them: 1) Epilepsy & Seizure Disorders, and 2) Neurobiology of Motor Maps. My work uses animal models of diseases, syndromes and treatments. I employ electrophysiological (Intracortical microstimulation, EEG, evoked potentials, multi- and single-unit activity, patch-clamp electrophysiology), and pharmacological (systemic and focal infusions) techniques to elucidate the underlying mechanisms. In collaboration with a number of colleagues I have also examined alterations in neural and glial cell anatomy at the cellular, ultrastructural and molecular/genetic levels. I measure animal behaviour using a variety of assessment techniques (e.g. open field, single pellet reaching task, pasta matrix task, elevated plus maze, cylinder task, and ladder task). I also use behavioural experience (skilled-reaching, wheel running, enriched environments) to understand how the brain changes.
Epilepsy and Seizure Disorder Studies
I am interested in discovering the relationship between seizures, behavioural disruption and their mechanistic underpinnings as well as methods to prevent or reverse seizure-induced alterations in behaviour and brain.
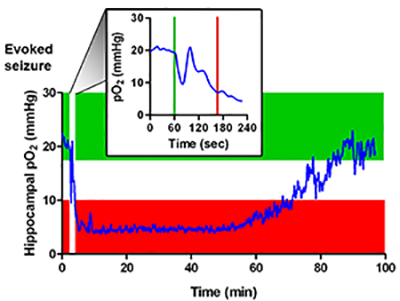
My laboratory has discovered that after a seizure terminates the brain has a stroke! See Farrell et al., 2016 eLife.
This event is caused by vasoconstriction leading to reduced blood flow and low oxygen levels and is characterized by hypoperfusion/hypoxia which is causally related to memory and motor disturbances. We have also discovered that the enzyme cyclooygenase-2 and L-type calcium channels play mechanistic roles in the expression of the postictal hypoperfusion/hypoxia. Our intention is to further interrogate the phenomenon, determine effective drugs that ameliorate the physiological underpinnings and behavioural expression and ultimately translate our findings to the clinic.
The postictal (after seizure) hypoperfusion/hypoxia event not only is responsible for acute dysruption of brain function but likely also plays a role in several other epilepsy related alterations in brain anatomy and function. For instance it may be that repeated hypoperfusion/hypoxia is responsible for chronic changes in brain anatomy (sclerosis and damage), astrocyte proliferation, blood-brain barrier breakdown, and increases in central inflammation. Moreover the behavioural comorbidities (like anxiety and depression) which are often observed in people with epilepsy could be caused by the hypoperfusion/hypoxia. Sudden Unexpected Death in Epilepsy (SUDEP) may be the result of vasoconstriction and low oxygen leves in brainstem nuclie responsible for breathing.
Neurobiology of Motor Map Studies
A long-standing and central question in Psychology and Behavioural Neuroscience is, “How does experience change the brain”? Many of us working on this vexing question believe that an alteration in the connections (synapses) between neurons allows a restructuring of neural circuits (engrams) to encode memories. Thus, the theoretical framework in which I work is aptly called the synaptic plasticity and memory theory. This theory also holds that a particular memory is distributed within the brain regions responsible for that behaviour and it is located among other previously formed memories.
My students, collaborators and I have published many experiments examining synaptic plasticity and movement representations (motor maps) within the rat forelimb area of sensorimotor neocortex. This area shows synaptic potentiation and depression phenomena that are correlated with anatomical changes. Most importantly, learning skilled behaviours of the forelimb are dependent on the integrity of this area of motor cortex and result in functional and anatomical changes. Thus, we have an easily accessible neuroanatomical region within which to focus our exploration of the relationship between brain and behaviour, as well as the reciprocal relationship between behaviour and brain.